Systems Biology - A Brief Summary
My Three Parts of Systems Biology in Hopefully "Simpler" Terms

I know that these three articles are difficult to understand and very theoretical in nature. That’s why I’ll summarize them briefly and as simply as possible:
Part 1 shows that local linearization cannot solve the trajectory problem.

By its very definition, a phase space is not a physical location but a mathematical one: every possible state of a biological system, defined simultaneously by all its variables (protein concentrations, membrane potentials, receptor occupancy, metabolic fluxes), corresponds to a single point in this space. Nevertheless, from a strictly systems biology perspective, it is the boundary within which systemic trajectories can operate. The hierarchy here is thus: membrane for the cell, tissue for cell assemblies, organ for tissue assemblies, and skin and bone for organs. This is necessary to organize a restriction and logic within the trajectories and to establish functionality.
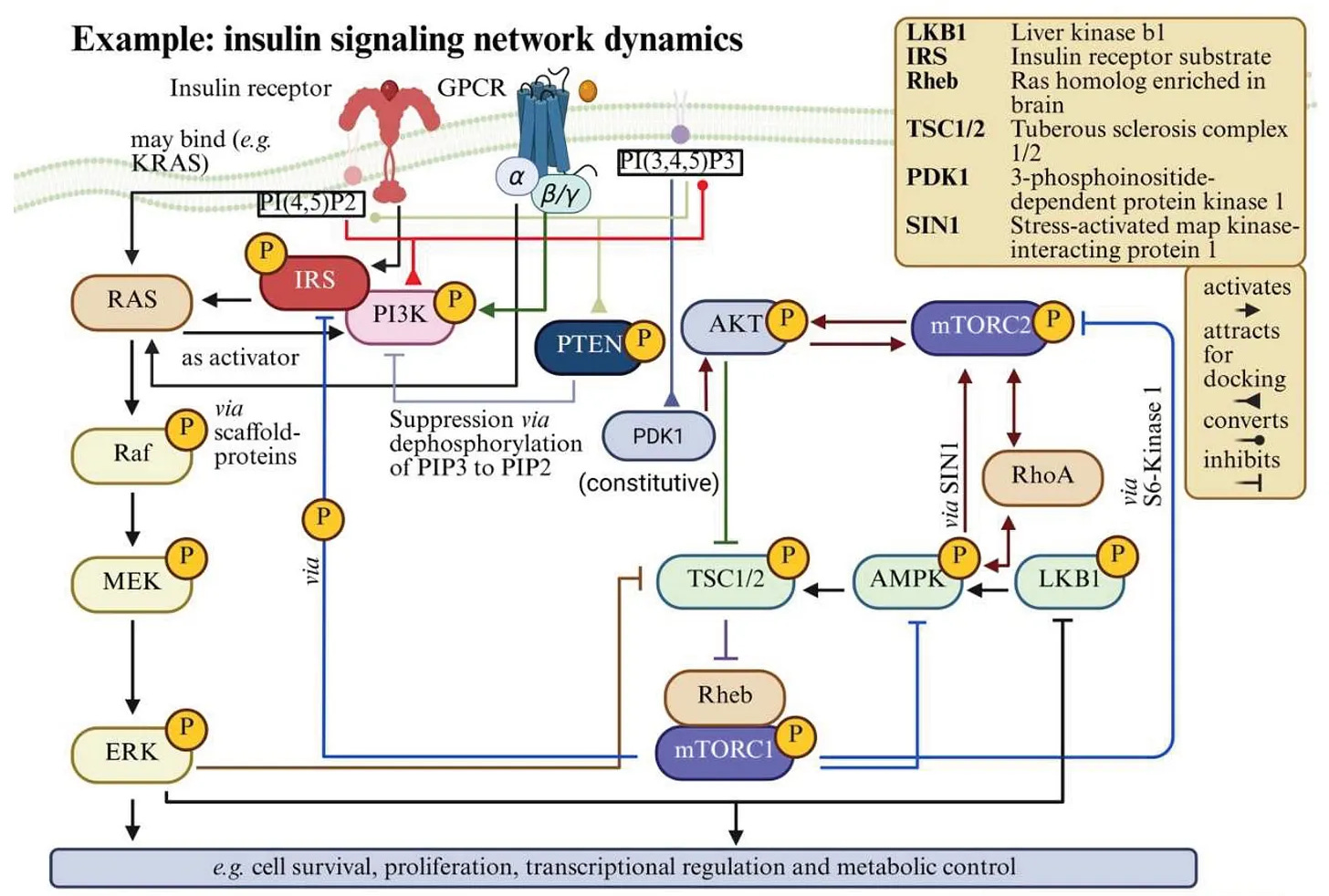
A trajectory is then the path a system traces through that space over time, the actual sequence of states it moves through. The Jacobi matrix, a standard tool in network analysis, describes how sensitive each variable is to small changes in the others, but only at one specific point. It is a local snapshot of the system's wiring at a given moment. The problem is that biological systems do not stay near fixed points: they oscillate, pulse, bifurcate, and switch between qualitatively different dynamic regimes. Once the system moves far enough from the reference state (as it does during signaling cascades, cell state transitions, or, critically, when an exogenous perturbation like an LNP alters the membrane environment) the local linearization no longer holds. The map is accurate only where you are standing, not where the system is going.
Part 2 shows that population data and reconstruction methods cannot solve it either, and that the problem scales upward, which means to the point where a higher-level coordination space becomes operationally necessary.

Even if you cannot measure a single cell's trajectory directly, you might hope to reconstruct it statistically from many cells at once. This is the logic behind single cell omics: sample enough cells, and the population distribution approximates the paths individual cells travel through gene expression space. But this logic has a structural flaw. As Hanin demonstrates, even small deviations from the assumption that all sampled cells are drawn from the same underlying distribution do not produce small errors; they produce systematic and irremovable bias. The cells are not replicates of one another. Each one occupies a distinct position in its own phase space, shaped by its epigenetic state, its signaling history, its local microenvironment. Averaging across them does not recover the trajectory; it erases it. And this problem does not stay at the cellular level. As organizational complexity increases, subsystems such as tissues, organs, and neural networks become coupled in ways that make each one dependent on the boundary conditions set by the level above. No subsystem can be fully understood in isolation, because its dynamics are continuously constrained by a higher dimensional context it does not internally contain. At some point, the coordination of these nested open systems requires what I call an operationally necessary space of movement: not a metaphysical addition, but a structural precondition for the system to remain coherent across its own hierarchy.
Part 3 formally demonstrates why this is the case: because every autopoietic system constitutes its own phase space from within and thus, in principle, undermines any external scaling logic.

Part 3 provides the formal answer to why external reconstruction and population averaging fail at a principled level, not merely a technical one. A living cell is not a passive object that occupies a predefined state space the way a molecule occupies a binding pocket. It actively constitutes the very dimensions of its own state space through the circular organization of its internal processes: the products of metabolism catalyze the reactions that produce those products. This is what autopoiesis means in operational terms. The system is closed to efficient causation, meaning no external agent directs its internal logic, while remaining materially open to energy and matter, and informationally open to formal constraints encoded in structures like DNA. These three properties coexist simultaneously, and their coexistence is not a paradox but the defining feature of living organization.
The LNP strand is not a separate topic woven into systems biology. It is the test case in which the thesis receives its sharpest physical formulation: If LNPs not only damage a membrane but also disrupt the physical conditions under which organizational closure is possible, then they constitute an intervention in what constitutes the system as a system. This is a different statement than “LNPs are cytotoxic”. Although it is harder to grasp, it ist the much more precise perspective:
The consequence for pharmacology is direct: when a drug or delivery vehicle such as a lipid nanoparticle interacts with a cell, it does not perturb a static target in a fixed landscape. It perturbs the physical conditions under which the cell constitutes its own organizational closure in the first place. Disrupting the membrane is not simply damaging a barrier; it potentially interferes with the topological self-generation that makes the entire causal architecture of the cell possible. This is why no fractal scaling model, no population average, and no local linearization can fully capture what happens, because the system being described is, by its own nature, self-describing, and that capacity is precisely what external frameworks cannot absorb.
Ahhhhh I'm getting there. Thank you.